| Dokdonia donghaensis | |
|---|---|
| Scientific classification | |
| Domain: | Bacteria |
| Phylum: | Bacteroidota |
| Order: | Flavobacteriia |
| Suborder: | Flavobacteriales |
| Family: | Flavobacteriaceae |
| Genus: | Dokdonia |
| Species: | D. donghaensis |
| Binomial name | |
| Dokdonia donghaensis Yoon et al. 2005 | |
Dokdonia donghaensis is a strictly aerobic, gram-negative, phototrophic bacterium that thrives in marine environments. The organism can grow at a broad range of temperatures on seawater media. It has the ability to form biofilms, which increases the organism's resistance to antimicrobial agents, such as tetracycline.
Discovery and isolation
Dokdonia donghaensis was discovered and isolated from the Sea of Japan in Korea from an island called Dokdo, a Korean name for the Liancourt Rocks which sovereignty is disputed between Japan and Korea; the date of isolation is not specified. Phylogenetic analysis of the 16S ribosomal RNA gene sequence of the organism revealed two closely related strains, DSW-1T and DSW-21. These strains are classified in Cytophaga-Flavobacterium-Bacteroides (CFB) group, which are dominant genera in marine environments. Winogradsky first described Cytophaga-like bacteria as unicellular, gliding, non-spore-formers, and Gram-negative rods, although the shape varies. Many Cytophaga-like bacterial colonies have pigment due to flexirubin-type pigments that are only found in these bacteria and flavobacteria. Cytophaga–Flavobacteria are chemoorganotrophs and are able to degrade biopolymers like chitin and cellulose.
Morphology and genome
D. donghaensis is a gram-negative bacterium that is rod-shaped, nonmotile, and a non-spore-former. The bacteria are able to grow on seawater media with agar, in which the colonies appear circular, slightly convex, glistening, smooth, yellow, and 1–2 mm in diameter. The DSW-1T strain of D. donghaensis has about 3,923,666 base pairs in its genome. The DNA G+C content for D. donghaensis is 38%.
Image of the phylogenetic tree can be viewed here.
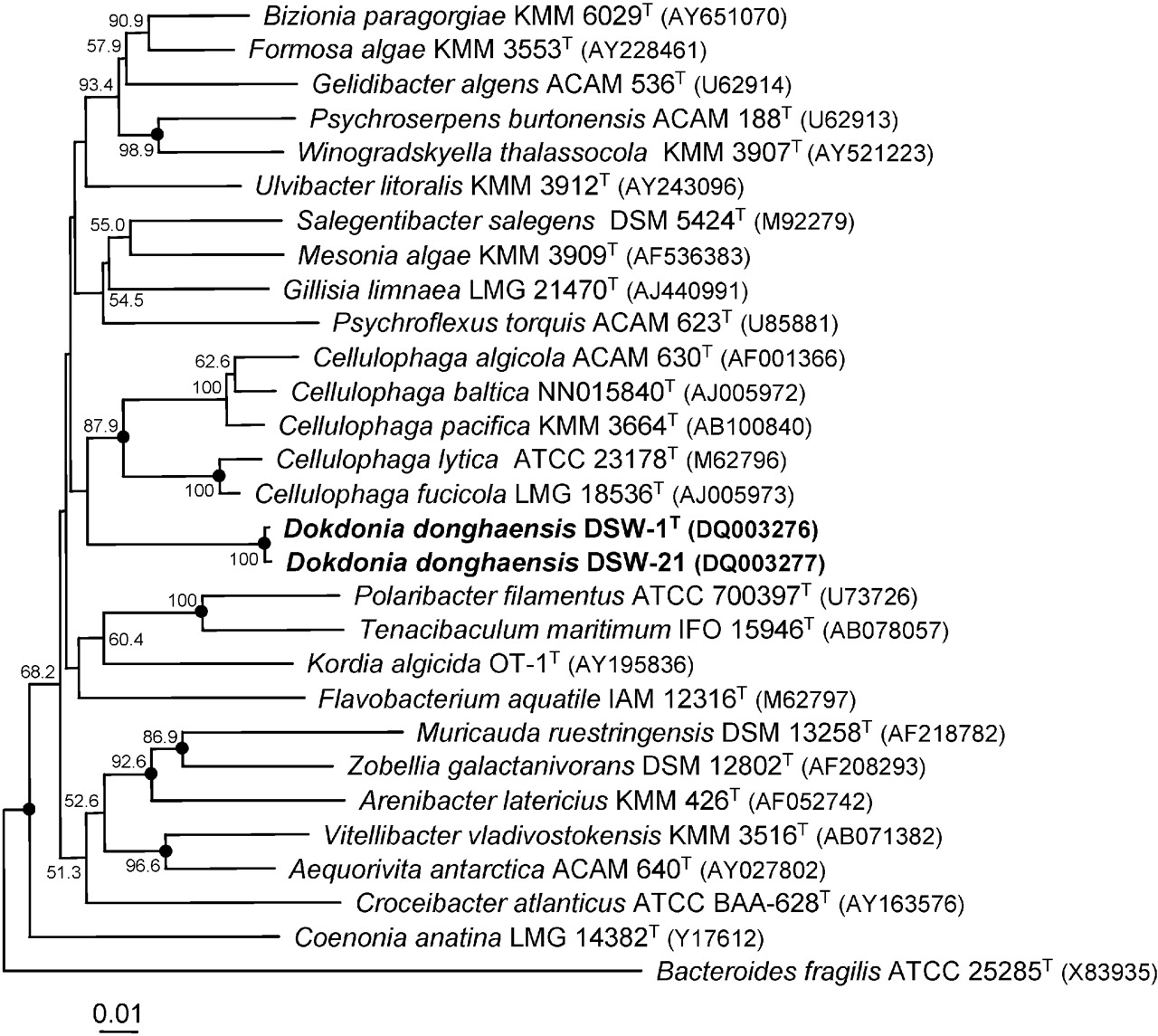{kind=link}
Pathology
D. donghaensis has not been identified as a human pathogen.
Metabolism
D. donghaensis performs light-driven sodium ion transport. This light-driven proton pump is called a proteorhodopsin, which increases the organism's survival rate because it can convert light into energy that the organism needs to grow. Since the organism uses light for energy, it is a phototroph. D. donghaensis is able to secrete enzymes such as catalase and oxidase. Since the organism is a strict aerobe, it used oxygen as a terminal electron acceptor. The organism can grow on peptone and tryptone as the sole carbon and nitrogen sources. D. donghaensis cannot grow in the absence of NaCl or when it is greater than 7% (w/v); growth is optimal at 2% (w/v) NaCl. The organism is susceptible to the antibiotics tetracycline and carbenicillin, although the organism's ability to form biofilms makes it difficult for antibiotics to penetrate the viscous layers.
Habitat
D. donghaensis lives in halophilic marine environments. The organism can grow as low as 4 °C and as high as 35 °C, although the optimum temperature for growth is 30 °C. The optimal pH for growth is 7–8, although growth can be observed at a pH as low as 5.5.
Biofilm formation
D. donghaensis is able to form biofilms in marine habitats, which is a survival strategy that allows the organism to grow while being protected from environmental stresses. Biofilm formation serves a purpose for marine bacteria in that it increases their resistance to antimicrobial agents, desiccation, and grazing. Biofilms allow the microbes to attach to surfaces by excreting extracellular polymeric substances (EPS). Marine bacteria that adhere to surfaces form host-specific and spatially structured communities that are fairly stable.
References
- "Dokdonia". www.uniprot.org.
- Parte, A.C. "Dokdonia". LPSN.
- Yoon, JH; Kang, SJ; Lee, CH; Oh, TK (2005). "Dokdonia donghaensis gen. nov., sp. nov., isolated from sea water". International Journal of Systematic and Evolutionary Microbiology. 55 (6): 2323–2328. doi:10.1099/ijs.0.63817-0. PMID 16280490.
- Bowman, JP; McCammon, SA; Brown, MV; Nichols, DS; McMeekin, TA (1997). "Diversity and association of psychrophilic bacteria in Antarctic sea ice". Appl Environ Microbiol. 63 (8): 3068–3078. Bibcode:1997ApEnM..63.3068B. doi:10.1128/AEM.63.8.3068-3078.1997. PMC 168604. PMID 9251193.
- Glöckner, FO; Fuchs, BM; Amann, R (1999). "Bacterioplankton compositions of lakes and oceans: a first comparison based on fluorescence in situ hybridization". Appl Environ Microbiol. 65 (8): 3721–3726. Bibcode:1999ApEnM..65.3721G. doi:10.1128/AEM.65.8.3721-3726.1999. PMC 91558. PMID 10427073.
- Kirchman, DL (2002). "The ecology of Cytophaga-Flavobacteria in aquatic environments". FEMS Microbiol Ecol. 39 (2): 91–100. doi:10.1016/s0168-6496(01)00206-9. PMID 19709188.
- Kim, K; Kwon, SK; Yoon, JH; Kim, JF (2016). "Complete genome sequence of the proteorhodopsin-containing marine flavobacterium Dokdonia donghaensis DSW-1T, isolated from seawater off Dokdo in the East Sea (Sea of Korea)". Genome Announc. 4 (4): e00804-16. doi:10.1128/genomea.00804-16. PMC 4974333. PMID 27491981.
- DeLong, EF; Beja, O (2010). "The light-driven proton pump proteorhodopsin enhances bacterial survival during tough times". PLOS Biol. 8 (4): e1000359. doi:10.1371/journal.pbio.1000359. PMC 2860490. PMID 20436957.
- Wang, Y; Zhou, C; Ming, H; Kang, J; Chen, H; Jing, C; Feng, H; Chang, Y; Guo, Z; Wang, L (2016). "Pseudofulvibacter marinus sp. nov., isolated from seawater". Int J Syst Evol Microbiol. 66 (3): 1301–1305. doi:10.1099/ijsem.0.000879. PMID 26739348.
- Burmølle, M; Webb, JS; Rao, D; Hansen, LH; Sørensen, SJ; Kjelleberg, S (2006). "Enhanced biofilm formation and increased resistance to antimicrobial agents and bacterial invasion are caused by synergistic interactions in multispecies biofilms". Appl Environ Microbiol. 72 (6): 3916–3923. Bibcode:2006ApEnM..72.3916B. doi:10.1128/aem.03022-05. PMC 1489630. PMID 16751497.
- Yoon, JH; Kang, KH; Park, YH (2003). "Psychrobacter jeotgali sp. nov., isolated from jeotgal, a traditional Korean fermented seafood". Int J Syst Evol Microbiol. 53 (2): 449–454. doi:10.1099/ijs.0.02242-0. PMID 12710611.
- Fux, CA; Costerton, JW; Stewart, PS; Stoodley, P (2005). "Survival strategies of infectious biofilms". Trends Microbiol. 13 (1): 34–40. doi:10.1016/j.tim.2004.11.010. PMID 15639630. S2CID 10216159.
- Mah, TF; O'Toole, GA (2001). "Mechanisms of biofilm resistance to antimicrobial agents". Trends Microbiol. 9 (1): 34–39. doi:10.1016/s0966-842x(00)01913-2. PMID 11166241.
- Jefferson, KK (2004). "What drives bacteria to produce a biofilm?". FEMS Microbiol Lett. 236 (2): 163–173. doi:10.1016/j.femsle.2004.06.005. PMID 15251193.
- Matz, C; Kjelleberg, S (2005). "Off the hook—how bacteria survive protozoan grazing". Trends Microbiol. 13 (7): 302–307. doi:10.1016/j.tim.2005.05.009. PMID 15935676.
- Pasmore, M; Costerton, JW (2003). "Biofilms, bacterial signaling, and their ties to marine biology". J Ind Microbiol Biotechnol. 30 (7): 407–413. doi:10.1007/s10295-003-0069-6. PMID 12884126. S2CID 1458649.
- Taylor, MW; Schupp, PJ; Dahllöf, I; Kjelleberg, S; Steinberg, PD (2004). "Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity". Environ Microbiol. 6 (2): 121–130. doi:10.1046/j.1462-2920.2003.00545.x. PMID 14756877.
- Bhadury, P; Wright, PC (2004). "Exploitation of marine algae: biogenic compounds for potential antifouling applications". Planta. 219 (4): 561–78. doi:10.1007/s00425-004-1307-5. PMID 15221382. S2CID 34172675.
External links
| Taxon identifiers | |
|---|---|
| Dokdonia donghaensis | |